

IINSIGHTS
Take a minute to write an introduction that is short, sweet, and to the point.

Article: Type I interferons induce an epigenetically distinct memory B cell subset in chronic viral infection.
-
• scATAC-seq and scRNA-seq reveal expansion of an MBC subset in chronic LCMV infection
• A distinct chronic MBC subset is associated with increased IFN-I-associated ISG signature
• The chromatin landscape of MBCs is established during a critical window early in infection
• IFN-I dynamics govern memory B cell epigenome and phenotype in chronic viral infection
Summary

Article: Sustained type I interferon signaling after human immunodeficiency virus type 1 infection of human iPSC derived microglia and cerebral organoids
-
• HIV-1 productively infects iPSC-derived microglia and triggers inflammatory activation
• HIV-1 infection of microglia results in sustained type I interferon signaling
• Type I interferon signaling persists in HIV-1-infected sliced cerebral organoids
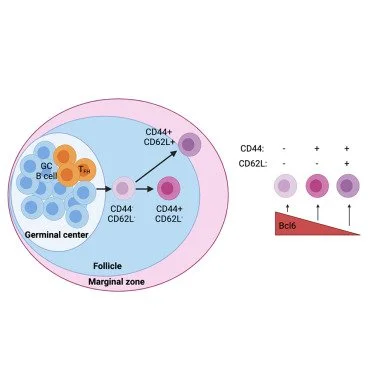
Article: CD62L expression marks a functionally distinct subset of memory B cells
-
• CD44 and CD62L identify transcriptionally and functionally distinct memory B cell subsets
• Bcl6 regulates memory B cell differentiation through control of a network of genes
• Overexpression of Zeb2 restricts the differentiation of CD62L+ memory B cells
• CD62L identifies phenotypically distinct human memory B cells following mRNA vaccination
Industry News and Articles

Article: Cancer-associated fibroblast phenotypes are associated with patient outcome in non-small cell lung cancer
-
• Imaging mass cytometry analysis of CAF heterogeneity in over 1,000 patients
• Definition of good and poor prognostic CAF phenotypes
• Poor prognostic CAFs are associated with metastasis and chemoresistance
• CAF spatial distribution correlates with immune infiltration and prognosis
Article: Sustained type I interferon signaling after human immunodeficiency virus type 1 infection of human iPSC derived microglia and cerebral organoids
-
• HIV-1 productively infects iPSC-derived microglia and triggers inflammatory activation
• HIV-1 infection of microglia results in sustained type I interferon signaling
• Type I interferon signaling persists in HIV-1-infected sliced cerebral organoids
Article: CD62L expression marks a functionally distinct subset of memory B cells
-
• CD44 and CD62L identify transcriptionally and functionally distinct memory B cell subsets
• Bcl6 regulates memory B cell differentiation through control of a network of genes
• Overexpression of Zeb2 restricts the differentiation of CD62L+ memory B cells
• CD62L identifies phenotypically distinct human memory B cells following mRNA vaccination
All-in-one IQ toggle switches with high versatilities for fine-tuning of transgene expression in mammalian cells and tissues
Jeongkwan Hong 1 6, Kyung-Cheol Sohn 2 6, Hye-Won Park 3 6, Hyoeun Jeon 3, Eunjin Ju 2, Jae-Geun Lee 4, Jeong-Soo Lee 4 5, Jaerang Rho 3, Gang Min Hur 2, Hyunju Ro 1
The transgene toggling device is recognized as a powerful tool for gene- and cell-based biological research and precision medicine. However, many of these devices often operate in binary mode, exhibit unacceptable leakiness, suffer from transgene silencing, show cytotoxicity, and have low potency. Here, we present a novel transgene switch, SIQ, wherein all the elements for gene toggling are packed into a single vector. SIQ has superior potency in inducing transgene expression in response to tebufenozide compared with the Gal4/UAS system, while completely avoiding transgene leakiness. Additionally, the ease and versatility of SIQ make it possible with a single construct to perform transient transfection, establish stable cell lines by targeting a predetermined genomic locus, and simultaneously produce adenovirus for transduction into cells and mammalian tissues. Furthermore, we integrated a cumate switch into SIQ, called SIQmate, to operate a Boolean AND logic gate, enabling swift toggling-off of the transgene after the removal of chemical inducers, tebufenozide and cumate. Both SIQ and SIQmate offer precise transgene toggling, making them adjustable for various researches, including synthetic biology, genome engineering, and therapeutics.
Graphical abstract
Keywords
transgene toggling
SIQ
tebufenozide
cumate
SIQmate
adenovirus
phiC31
Introduction
Tight manipulation of transgene induction, governed by a sophisticated engineered promoter with regulatable enhancer elements, has shed light on new avenues for elucidating the physiological function of genes of interest and for clinical applications of gene therapy. Since the application of the lac operator-repressor1 and tetracycline (Tet)-responsive transgene activation system2 on mammalian cells, tremendous efforts have been devoted to developing far more reliable toggle gene switches applicable not only at the cellular level, but also at the whole organism level.3,4,5,6,7 Thus far, developed transgene switches exploit tunable external stimuli, for instance, discrete hormones, chemicals, heavy metals, or physical stresses such as heat, light, magnetic fields, ultrasonic sound, and electrical signals.8,9 Among the aforementioned stimuli, chemically based systems have been highly sought because of the convenience of manipulating transgene induction levels via simple dosing with a small molecule at different concentrations and for different time windows.
Among the small molecule-dependent transgene switches, the Tet transactivator (Tet-On/Off) systems, exploiting Tet or its derivatives like doxycycline and minocycline as a triggering cue, and the Gal4/UAS system adopting upstream activation sequence (UAS) and a chimeric protein composed of Gal4 DNA binding domain (DBD), small molecule binding domain (ligand-binding domain), and transactivator domain (AD) responsive to appropriate chemical inducers such as RU486 (mifepristone) or tebufenozide (Teb), have been most widely used to modulate transgene induction and scrutinized for clinical applications.6,10,11,12,13,14 However, although exploiting the Tet-On/Off system has been considered as the most powerful method to induce transgene expression, the frequently observed unacceptable level of leakiness constrains its broad applications in biological researches, especially when tight control of transgene expression is required. The flaw of the Tet-On/Off system was significantly rectified by consolidating the binary mode of the Tet-On/Off system into a single vector optimized for lentivirus production.15,16,17,18 The all-in-one Tet-Off system decreased basal leakiness, but at the expense of the strength of the transgene induction ability.15 Despite several improvements applied to lentiviral-based transgene switches, the intrinsic propensity of its host genome integration must be considered when applying the lentiviral switch to gene therapy.18 While nonintegrating lentiviral vectors (NILs) have been developed19,20 and used for gene editing with a singular mode,21 chemically elicitable singular transgene switches based on NILs have not been reported yet. Meanwhile, the critical flaw of the Gal4/UAS system is its weaker potency in inducing transgene expression compared with other gene switches, such as the Tet-On/Off system.14 In a preclinical approach, a Gal4/UAS transgene induction system had been successfully delivered into the rat brain via adeno-associated virus (AAV), which was able to be stimulated by the intraperitoneal injection of RU486.22 The binary mode of the Gal4/UAS-based AAV transgene switch was ingeniously consolidated into a single AAV vector in the following studies.23,24 In these studies, regulatable glial cell-derived neurotrophic factor expression in the rodent brain by boosting the function of RU486 has shown therapeutic potential by ameliorating the symptoms of neurodegenerative disorders. Thus, the ideal transgene toggle switch should possess the singular qualities of being non-leaky while maintaining strong transgene induction potency.
A repressible binary transgene induction system, the Q switch, derived from the fungus Neurospora crassa,25 was adopted in various organisms as a novel transgene induction system.26,27,28,29,30,31,32,33 After extensive modification by several research groups, the Q switch became one of the most attractive transgene switches, with various beneficial features, including exquisite sensitivity, relatively low toxicity, and being free from transgene silencing problems. Recently, we reported a more reliable binary Q switch, IQ-Switch, optimized for the transgenesis of zebrafish.34 The IQ-Switch showed almost undetectable leakiness of transgene expression, eliciting at least 4.5-fold more sensitivity to a chemical inducer Teb in comparison to Gal4/UAS under identical experimental conditions in zebrafish.34
We previously demonstrated that the Gal4/UAS-based singular gene switch (EUI) was successfully implemented for transgene induction in human cell lines and rodent tissues.14,35 However, there are several issues that need to be critically improved for broader usability in biological research. First, the transgene inducibility of the Gal4/UAS system is generally weaker than that of other widely used gene switches, such as the Tet-On/Off system. Second, the Gal4/UAS system is highly vulnerable to enhancer methylation, which can result in transgene silencing after consecutive generations. Third, since there are two discrete vectors optimized for transient transfection (pEUI(+)) and adenovirus production (pENTR-EUI), respectively, combining the two features into one would be ideal for the ease of conducting experiments. Fourth, establishing stable cell lines using pEUI(+) has not been an easy task because the vector lacks any auxiliary sequences that aid in its genome integration.
Here, we report a novel singular IQ-Switch (henceforth SIQ) equipped with all the necessary components for transgene induction in mammalian cells and rodents’ tissue. The SIQ shows none of the leakiness of the transgene expression while maintaining high sensitivity to the chemical inducer. We also included two distinct integrase recognition sites to induce genome integration into a specific locus and to produce adenoviral vectors via recombination events. With a single vector of SIQ, we successfully generated several cell lines with switchable transgenes integrated at an identical genomic locus and efficiently produced adenovirus for the transduction of the same tunable transgenes into cells and tissues that are refractory to conventional transfection approaches. Another advantage of the singular feature of SIQ is that no further molecular cloning is required for adenovirus preparation. In addition, by adapting transcriptional Boolean logic gates, we tested the utility of SIQ for synthetic biology purposes. We showed that the SIQ-based logic circuit combined with a transcriptional repression device guides more swift togging-off of transgene switch. Altogether, the novel SIQ and its derivative enable the researchers to investigate the cellular roles of genes of interest under physiologically relevant conditions with tunability, robustness, versatility and reliability.
Results
The “all-in-one” SIQ is far more potent for transgene induction than Gal4/UAS
To address the flaws of EUI gene switch, we attempted to integrate the binary IQ-Switch, originally optimized for zebrafish transgenesis,34 with several modifications. To create a novel SIQ, we assembled all the necessary elements for IQ-based transgene toggling into a pENTR-EUI vector. The Gal4-VP16-EcR transactivator of pENTR-EUI was replaced with an IQ-Switch module composed of a QFDBD (which recognizes QUAS), a 2× QF minimal activation domain (2×AD∗), a minimal VP16 activation domain (VP16∗), and a domain of the ecdysone receptor (EcR), all of which are located upstream of a bGH poly(A) signaling sequence. We substituted 13×QUAS for the 10×UAS of the vector, followed by an SV40 poly(A) signaling sequence. The multiple cloning site (MCS) located immediately after a carp beta-actin minimal promoter consisted of the following restriction enzymes in sequence: BspD I, Cla I, BspE I, Afl II, Not I, and Sca I. To facilitate the establishment of cell lines with genes of interest, we introduced a phiC31 serine integrase recognition site, referred to as attB,36 downstream of a bGH poly(A) signaling sequence and upstream of the 13×QUAS. Additionally, a puromycin selectable marker, controlled by the SV40 promoter, was inserted between the pUC origin (ori) of replication and the attL1 recombination site. The kanamycin resistance gene remained unaltered. The finalized vector, designated as pSIQ (GenBank accession number: OR355468), was designed to lose both the pUC ori and two selectable markers after recombining with a pAd/PL-DEST (Invitrogen) vector for the preparation of adenovirus (Figures 1A and 1B).
Figure 1. A schematic diagram of SIQ transgene toggle switch
(A) The pSIQ vector encompasses all the components of an IQ-Switch, including an IQ-Driver: QFDBD, two minimal activation domains of QF (2×AD∗), a minimal VP16 activation domain (VP16∗), and an EcR. These components are governed by a CMV promoter and required for binding to the 13× QUAS enhancer elements, subsequently inducing transcription of genes of interest only when stimulated by Teb. A phiC31 serine integrase recombination site is denoted as attB, where the recombination event occurs to the corresponding attP site in the presence of the recombinase. For ease of cloning into the pAd/PL-DEST destination vector, we included gateway cloning-associated elements (attL1/attL2) for the production of adenovirus. The kanamycin and puromycin antibiotic-resistant markers are denoted as KanaR and PuroR, respectively. (B) The all-in-one SIQ transgene toggle switch exhibits high versatility. With a single construct, a broad range of tailored transgene induction experiments can be carried out through transient transfection, generation of stable cell lines, and adenovirus-mediated transgene delivery. The arrows indicate that discrete purposes of experiments could be merged into a SIQ.
To investigate whether pSIQ could operate as expected in human cell lines, we performed transient transfection in HEK293 cells using pEUI(+) for a direct comparison. Although both plasmids exhibited EGFP transgene expression in a Teb dosage-dependent manner, the expression from pSIQ was significantly more potent than that from pEUI(+), even when EGFP expressivity seemed to saturate at doses exceeding 5 μM of Teb (Figure 2A). The enhanced sensitivity of pSIQ compared with pEUI(+) was quantitatively validated using a dual luciferase assay. As shown in Figure 2B, in comparison with the inducibility of the EGFP transgene, both plasmids reached a plateau at 1 μM Teb. Notably, pSIQ stimulated the luciferase reporter significantly more than pEUI(+) did at any Teb dose. Importantly, we observed an approximately 40% lower basal level of leakiness with pSIQ compared with pEUI(+) (Figure 2C). Taken together, our data show that pSIQ is far more potent and reliable than the previously suggested pEUI(+) transgene toggling system.
Figure 2. SIQ surpasses the Gal4/UAS-based singular gene switch regarding sensitivity and the issue of leakiness
(A) The pEUI(+)-EGFP refers to a previously reported singular gene switch based on the Gal4/UAS system. The gradually elevated induction of the EGFP reporter, dependent on Teb dosage, was visualized under a fluorescence microscope after transiently transfecting 1 μg of the indicated plasmid constructs. After 24 h of transfection, Teb was treated for 24 h, and then the expression level of EGFP was observed using the same exposure times and contrasts. Scale bar, 200 μm. (B) Quantitative measurements of transgene expression levels were performed using the dual-luciferase assay. HEK293 cells were transfected with either pEUI(+)-Luciferase or pSIQ-Luciferase. After transfection, the cells were treated with Teb for 8 h. Statistical significance was analyzed using two-way ANOVA with Šidák correction (n = 3). ns, not significant. (C) The basal leakiness of pEUI(+) and SIQ was compared through a dual luciferase assay in the absence of Teb. Statistical significance was explored using the Student t test. The data are represented as mean ± SD (n = 3). ∗∗∗p < 0.001. A.U., arbitrary unit; Luc, luciferase.
SIQ makes it easy to generate stable cell lines
The establishment of stable cell lines has many advantages, encompassing features for studying the exquisite mechanisms of genes or the development of new therapeutics.37 However, it is concomitant with laborious procedures. As we mentioned above, the pSIQ equipped with serine integrase recognition sequence (attB) provides a convenient method for generating stable cell lines through integrase-based genome recombination.36 To facilitate the genome integration of transgenes subcloned in pSIQ, we initially allocated an attP site into the genome of host HEK293 cells by means of a lentiviral transduction system. The commercially available lentiviral vector (pCDH-CMV-MCS-EF1α-Puro, System Biosciences) was subjected to modifications to replace the antibiotic-resistant gene, puromycin N-acetyltransferase, with blasticidin deaminase, whose expression is governed by an EF1α promoter. Additionally, the CMV7 promoter was removed, and an attP phiC31 recognition site was subcloned into the upstream of EF1α promoter. We used the vector designated as pBla-attP-Lenti for packaging lentivirus to infect HEK293 cells. After subjecting the cells to selective pressure with blasticidin S antibiotics, the surviving cells were individually selected to establish stable clones (HEK293-attP-Bla), each carrying the attP site. We co-transfected the pSIQ encoding an EGFP transgene with the phiC31 integrase expression vector in HEK293-attP-Bla. Subsequently, we treated the cells with puromycin to select those resistant to the antibiotic treatment. Successfully integrated clones (SIQ-EGFP) were then validated by genomic PCR (Figures 3A and 3B), confirming their correct implantation into the attP site of the host genome.
Figure 3. Stable cell line harboring the SIQ system could be generated by integrating it into a specific locus of the host cell genome
(A) A schematic diagram illustrating the phiC31-mediated genomic integration of the SIQ toggle switch. The phiC31 serine integrase targets the attB element of pSIQ and an attP site in the host cell genome, mediating directional recombination, resulting in producing attR and attL recombined sequences. (B) A representative result of genomic PCR using the primers depicted with red and black arrows in (A). The first PCR products amplified by primers labeled with red arrows were used for the second PCR reaction with primers depicted by black arrows. The expected size of PCR products was shown in (A) under the square brackets. c, control group with mock cell genome for PCR template; e, experimental group. (C) Established stable cell lines harboring SIQ-EGFP were exquisitely sensitive to Teb stimuli. EGFP was detected only in the group of cells treated with 10 μM Teb for 24 h. Bright field images were located on the left side. Scale bar, 200 μm. (D) EGFP was detected as early as 3 h after 10 μM of Teb treatment, as shown by western blotting. Endogenous expression of α-tubulin was used as a loading control. (E) After 24 h of Teb (10 μM) treatment, the cells were reseeded into fresh culture dishes to minimize Teb presence. The level of EGFP transcripts was measured by qRT-PCR after harvesting the cells at the indicated time points. The data are presented as mean ± SD. Scale bar, 200 μm. Statistical analysis was performed using One-way ANOVA with Tukey’s HSD test (n = 3). ∗∗∗p < 0.001. (F) Schematic comparison between pSIQ-EGFP without IQ-Driver (W/O Driver) and pSIQ-EGFP constructs. (G) FACS analysis of Mock (containing only an attP in their genome), SIQ-EGFP W/O Driver, and SIQ-EGFP stable cell lines after 24-h exposure to Teb (10 μM). Data are presented as mean ± SD (n = 3).
To induce EGFP expression, we treated the SIQ-EGFP with Teb. As shown in Figure 3C, the EGFP fluorescence signal was exclusively detected in cells treated with Teb. After Teb stimulation, we observed a rapid increase in EGFP expression, detectable as early as 3 h after treatment (Figure 3D). The expression level of EGFP continued to rise with continuous Teb treatment, reaching a plateau at 48 h (Figure 3D). Importantly, the established cell line responded exclusively to exogenous chemical stimuli, as evidenced by the gradual decrease in the level of EGFP transgene as time goes on until 48 h after the removal of Teb (Figure 3E). This ability of SIQ to fine-tune the EGFP expression suggests that the toggle switch could be a valuable tool for various cellular applications. To revalidate the high inducibility of Teb and the low leakiness in the SIQ system, fluorescence-activated cell sorting (FACS) analysis was performed. We aimed to demonstrate the fidelity of SIQ by generating another stable cell line with an EGFP transgene and all the SIQ components except for the driver module (CMV-QFDBD-2x AD∗-VP16∗-EcR-poly(A)). This cell line was targeted to the identical attP locus of HEK293-attP-Bla, enabling a direct comparison with SIQ-EGFP (Figure 3F). As expected, the cell line lacking the driver module exhibited no response to Teb stimuli, while over the 94% of SIQ-EGFP cells were sorted as EGFP positive when stimulated with Teb (Figure 3G). Importantly, the SIQ-EGFP without Teb treatment displayed only a negligible background level of EGFP expression (Figure 3G).